揭秘青鳉抗病毒感染:负调控因子jun的鉴定之旅



文 | 十一情报局
编辑 | 十一情报局

●—前言—●
宿主的原癌基因jun(Jun protooncogene,AP-l transcription factor subunit)是一类表达广泛、多功能的转录调节因子,不仅和癌症、血管生成息息相关,也参与了机体的免疫调节和病毒的感染过程。
Jun属于转录因子激活剂蛋白I(activa-ted protein 1,AP-1)的亚族,包含 3个重要的蛋白结构域:Delta 结构域、反式激活结构域和DNA结合与二聚化结构域叨。

研究发现JUN能被JNK(c-Jun N-terminal kinase)磷酸化激活,并与JUND(JunD proto-oncogene, AP-I transcription factorsubunit)在肾上腺储质素反应中二聚化形成 AP-1参与到肿瘤的进程中。
研究表明Jun参与病毒相关的免疫调节通路,例如,在牛痘病毒(vaccinia virus,VACV)侵染鼠胚胎纤维母细胞的实验中,病毒激活Jun 的表达,从而促进病毒早期复制,加速病毒的扩散。

凡纳滨对虾(Litopenaeus wammamei)jun对具有先天性免疫功能的血蓝蛋白的转录调节至关重要C,此外,Jun 能结合到凡纳滨对虾基质金属蛋白酶mmp-2( matrix melalloproteinase-2)启动子上,激活 mmp-2的表达,而mmp-2又与许多的抗菌活性相关。
在红唇胭脂鱼(Liza haematocheila)中,jun和os都能调节ap-的启动子进而对免疫反应发挥重要作用。

在斜带石斑鱼(Epimephe-[us coioides)的GS细胞实验中,过表达突变型jun后,病毒的转录水平降低的同时细胞病变效应出现延迟。
在禽类成纤维母细胞中,宿主和病毒编码的高致癌性蛋白IUN(V-IUN)因具备分子的同源性,成为病毒复制过程中的重要媒介。
以上信息表明,jun是宿主抗病毒信号通路中潜在的负反馈因子,可能是鱼类分子抗病育种中的一个候选因子,需要进一步进行验证。

日本青鳉 (Oryzias latipes)是一种生殖周期短、便于实验室养殖、具有公开的基因组、容易开展基因编辑育种的模式鱼类,对其基因的功能分析有助于养殖鱼类的基因功能验证。
为了进一步了解jum基因在鱼类抗病毒天然免疫方面的功能,本研究以日本青鳉为研究对象,采用 CRISPR/Cas9介导的基因编辑技术获得 jun缺失的纯合子突变体品系,并对其在神经坏死病毒《nervous necrosisvirus,NNV)感染过程中的特征展开研究,以期为养殖鱼类的抗病育种提供理论依据。


●—研究探索—●
日本青鳉为华中农业大学水产学院模式鱼类繁育实验室饲养的HdrR 品系,养殖在水温28℃光照周期明:暗为14 h: 10h的循环水系统中。
性成熟的青鳉通过自然繁殖的方式获得后代,胚胎放置于培养液中,在28 ℃培养箱育。

在NCBI上下载预测的青jun cDNA序列(gene ID:LOC101173011),设计扩增编码区(cod-ing sequence,CDS)的引物(表 1)。
选择表达量高的性腺,采用RNAiso Plus试剂盒提取总RNA,使用PrimeScript™RT Reagent Kit with gDNA Eraser反转录试剂盒(TaKaRa,日本)经过反转录获得CDNA。
采用Q5高保真的DNA聚合酶(Biolabs,美国)进行PCR扩增获得jun的CDS序列,反应条件为94 ℃3 min;35 个循环:94 ℃30s ,60℃30 s,72℃60 s:最后72 ℃延伸10 min。

随后将扩增的PCR产物进行1%琼脂糖凝胶电泳,确定大小正确后,回收目的片段连接到pMD18-T(TaKaRa,日本)载体,连接产物转化到 DH5a感受态细胞中,送阳性克隆至武汉擎科生物有限公司测序。
在NCBI网站下载人、鼠,猪、斑马鱼和青鳉的Jun和同家族Junb、Jund 的序列,通过MEGA7中的ClustalW 方法进行蛋白的多序列比对,使用Neighbor-joining方法构建系统进化树,Bootstrapmethod 参数设置为1000。

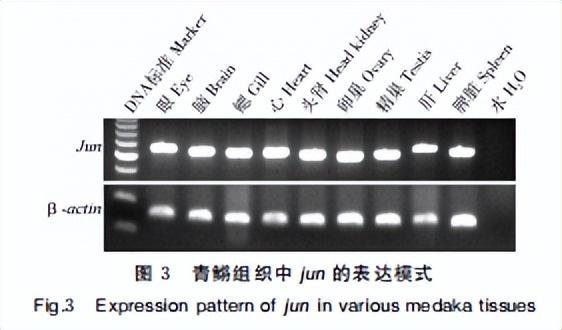
提取野生型青鳉成鱼的眼、脑、鳃、心脏、头 肾、卵巢、精巢、肝脏、脾脏9个组织的总RNA,反转录获得各个组织的cDNA。
设计引物jum RT-F/RT-R(表 1),通过半定量PCR检测基因在各个组织表达,以Bactin基因作为内参,PCR反应条件为:95℃变性30 s,60℃退火30 s,72 ℃,25个循环,延伸10 s,25个循环,PCR 产物在 2%琼脂糖凝胶上分离。

根据NCBI上的日本青鳉jun基因序列,在网站 https
://cctop.cos.uni-heidelberg. De: 8043/上设计合适的敞除靶位点,选择分值高且脱靶风险小的靶点jun sgl/sg2 F(表 1)。
在靶序列5’端前加T7启动子序列,在3’端加sgRNA scaffold引物扩增序列,具体的sgRNA(sigle guide RNA)的上游引物为:5’ TGTAATACGACTCACTATAGG-(ERNA20 nt)-GTTTTAGAGCTAGAAAT 3’,下游引物为来自模板PMD19T-sgRNA的通用引物gRNA-R(表1),引物由武汉警科生物技术有限公司合成。
以pMD19T-sgRNA为模板用PCR扩增出双链的sgRNA,反应条件如下:95℃ 30 s;95 ℃ 5s,60℃C 30 s,72℃10 s,34个循环,然后使用T7转录试剂盒(mMESSAGE mMACHINET™T7 tran-scription kit,Thermofisher)转录出sgRNA,并使用氯化理沉淀法纯化回收 sgRNA。
Cas9 mRNA按照文献纯化制备,将Cas9(300 ng/µL)和sgRNA(50 ng/µL)的混合液注射到1细胞期的青胚胎。

在胚胎发育4d后提取注射的部分胎DNA,使用检测引物jun DF/DR(表 1)PCR扩增jun的靶位点片段,突变型的PCR产物与野生型的相比明显变短。
将注射的FO胚胎养至成鱼,然后通过尾检测PCR产物来筛选FO突变成鱼将突变体FO代成鱼与野生型杂交,获得F1代突变体将F1代养至成鱼,再通过单克隆测序筛选出F代中突变一致的类型进行自交得到的F2代并养至成鱼,通过PCR筛选出代中的纯合子突变体,并将突变的纯合子繁殖扩大以供后续实验使用。
收取野生型青鳉胚胎,待仔鱼孵化出膜3d后每组15尾放人35 mm培养中加人总体积为4 mL的胚胎培养液,设置 0、2、10、50、100、200 µL神经坏死病毒进行病毒浓度梯度的预实验,以确定半致死病毒剂量。
实验中的NNV是从所在实验室从患病石斑鱼分离获得的,采用 RGNNV的保守引物经过PCR扩增全长后测序,序列确定为RGN-NV,其包含2条RNA链,分别为RNA(2.9 kb,NCBI序列号M2053461)和 RNA2(1.4 kb,NCBI序列号MN105076.1),将病毒在石斑鱼GE细胞中扩增后保存于-80 C(未发表)。

参考预实验数据对仔鱼进行病毒感染实验,试验设置空白对照组(野生青鳉仔鱼不加NNV)、对照组(野生青鳉仔鱼加NNV)和实验组(jum -/A124突变体仔鱼加NNV)。
对照组和实验组均加人100 pLNNV,每隔12h观察记录每组的死亡数,实验共重复7次,最后绘制对照组和处理组的死亡曲线。

对成鱼进行病毒感染实验时,使用微量注射器从野生型《对照组)和突变体成鱼《实验组)的泄殖孔向腹腔内注射 25 pLNNV,每组注射 10 尾,感染2周后取样。
由于NNV 主要攻击神经组织密集的脑和眼组织,提取脑、眼、鳃和肌肉4个组织RNA,通过qRT-PCR检测jun-/- A124和野生型各组织中NNV病毒的相对复制量(表 1)。
设计与干扰素通路相关的4个基因jun、fosl1(FOS-like antigen 1) ,tbk 1(TANK-binding kinase1) , irf3(interferon regulatory factor3)的qRTPCR 引物(表 1),检测在NNV病毒诱导下,jum-/-△124和野生型的个体中各基因的表达。
实验对象为孵化出膜3d的仔鱼,共设置4组。,即野生对照组(野生青鳉仔鱼不加NNV诱导)野生实验组(野生青鳉仔鱼加NNV诱导),突变体对照组《突变体青仔鱼不加NNV诱导)和突变体实验组(突变体青鳉仔鱼加NNV诱导)。
每组10尾NNV诱导剂量为100 pL,病毒诱导4 d后取样,用gRT-PCR检测4组样品中各基因的表达。

每组各取3尾鱼进行检测同时设置阴性空白对照,使用B-actin作为内参基因,基因的相对表达量采用 2~aC法进行计算。
试验数据均用"平均值±标准差”表示,并用GraphPad Prism 5件 t-tests中的Two-tailed对实验结果进行统计学分析,显著性差异设为 * P<0.05** P<0.01,* ** P<0.001,* *** P<0.000 1。

●—结果与分析—●
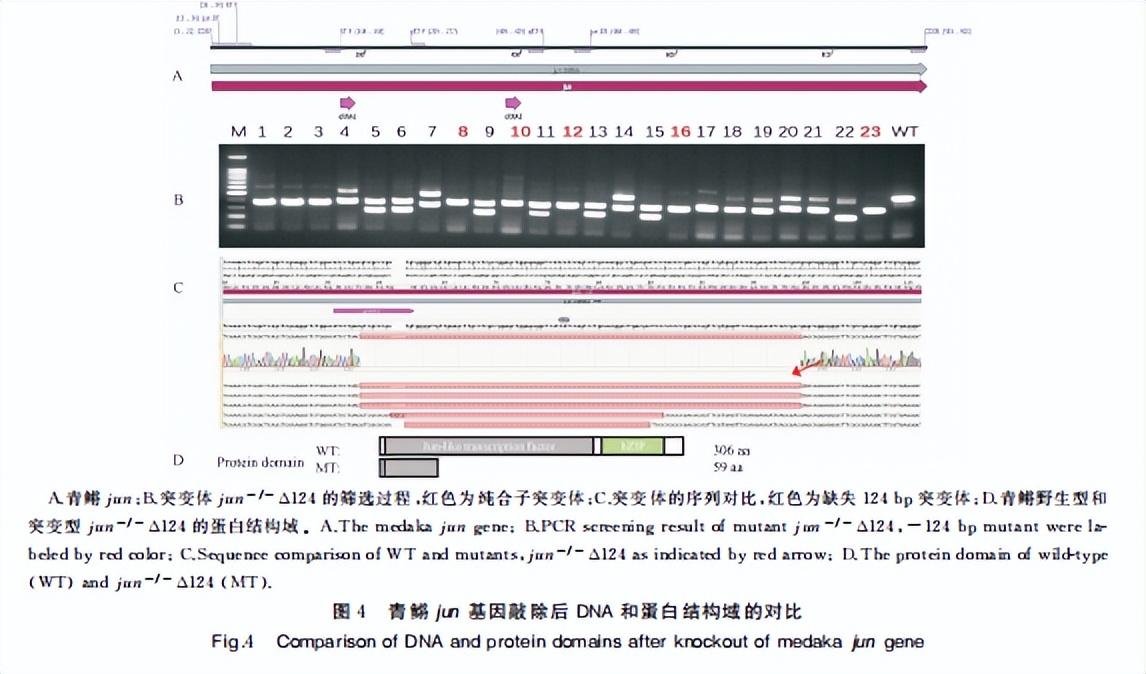
采用高保真DNA聚合酶,使用表1中的引物,PCR扩增后得到目的条带约为1 000 bp的jun CDS片段,克隆到T载体后,对目的条带进行测序。
测序获得的青鳉jun的CDS由起密码(ATG)到终止子(TAG)全长为921 bp,共编码306个氨基酸(aminoacid,aa),包括N端(5~209 aa)的Jun超家族结构域和C端(230~290 aa)的 bZIP 超家族结构域(图 1)。

通过系统进化分析可以看出,Jun、Junb和Jund在各支内都表现为硬骨鱼纲的青鳉和斑马鱼亲缘关系更近,而哺乳纲的人、鼠、猪亲缘关系更近(图 2)。

以Bactin为内参,对眼、脑、鳃、心脏、头肾、卵巢、精巢,肝脏、脾脏9个组织内jun的基因表达进行半定量PCR检测,发现其在所检测的9个组织中都表现出较高的表达水平,检测结果如图3所示。
通过显微注射将合成好的靶点 sgRNA1(5’ATCCTGACGTCGCCCGATGT3’)/SgRNA2(5’GGACAGACAGTTGCTACCGG3’)和Cas9 MRNA共注射到早期胚胎。
经过FO代突变体亲本筛选F代同种突变类型杂合子单克隆筛选和F2代纯合子突变体筛选,获得了jun缺失124个藏基的纯合子的突变体品系(MT:ju-/- △124)。
对该突变体品系的DNA和蛋白结构进行预测(图 4),发现突变体蛋白缺失了Jun家族结构域和bZIP结构域,在第59个氨基酸处出现翻译终止,为有效的突变类型。
根据最终获得的死亡曲线(图 5A)可以看出jun-/-△124突变体品系相对于野生型具有明显的抗病毒效果。
仔鱼死亡的判别依据:躯体明显腐坏呈骨架状或肿大,仔鱼眼睛明显肿大且眼间距增大仔鱼的尾部明显弯曲,蝶点消失(图 5B)。
另外,在成鱼的病毒感染实验中,在突变体的眼(图 5C)、脑(图 5D)、鳃(图 5E) 和肌肉(图 5F)中NNV的RNA2基因的表达明显低于野生对照组。
这说明jun-/-△124 突变体的NNV病毒量较少,因此突变体相较于野生型对 NNV 更为耐受。

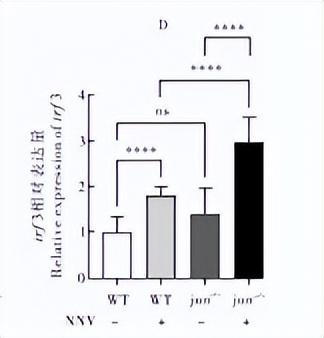
通过qRT-PCR检测干扰素相关基因jun、fosl1、tkkl、ixf3 在NNV诱导下的野生型和突变体内的表达情况,发现在病毒诱导后,野生型内的相关基因表达都有不同程度的上调,说明这些基因参与了抗病毒免疫反应。
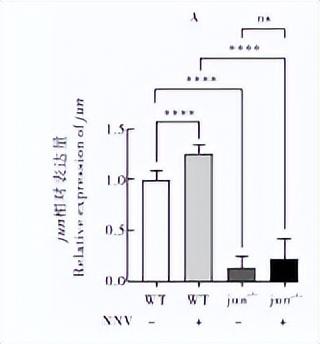
jun在jun -/- 124突变体中的表达极显著降低(P<0.000 1)(图 6A)说明jun的表达在突变体内受阻;在无病毒诱导情况下,jun-/-△124突变体中fosl1的表达相对于野生型有极显著增高(P<0.001),但在病毒诱导下相对于野生型的表达极显著降低(P<0.000 1),而在jun-/-△124突变体中fosl1的表达在病毒诱导或不诱导下均没有显著差异(图 6B)。
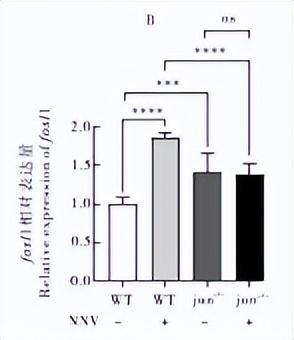
在无病毒诱导下,tkk1在jun-/-△124突变体中的表达相对于野生型没有显著差异,但在病毒诱导下相对于野生型也有降低(P<0.05),同时在jun-/-△124突变体中kk1 的表达在有无病毒诱导下两者也没有显著差异(图 6C)。
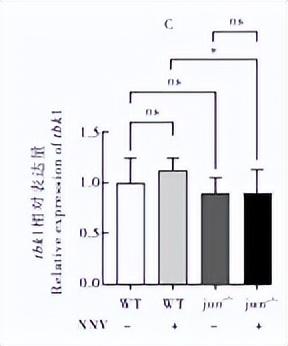
irf3 在不诱导的条件下,在jun-/-△124突变体中相对野生型没有显著差异,但是在病毒诱导的突变体内irf3的表达水平有极显著的上调(P<0.000 1),同时在突变体jun-/-△124中病毒诱导之后的表达量极显著升高(P<0.000 1)(图6D)。
宿主的jun是机体参与病毒感染和免疫调节的一个重要因子,本研究克隆了青鳉的jun 基因, 并通过基因编辑技术构建了突变体,初步研究发现其具有明显的抗病毒效果,而且突变体成鱼体内的病毒相对含量要比对照组显著降低,因此认为青鳉的jun 是病毒感染过程中的负反馈因子。

转录因子AP-1由许多的同源或异源的二聚体结合形成,包含 bZIP(basicregionleucinezipper, bZIP)转录因子家族结构域的成员,比如Jun、Fos、Atf。
研究发现,很多致癌性的病毒通过编码病毒bZIP转录因子来影响宿主细胞的免疫反应,使得病毒的复制更为有利。

在本研究中鉴定的青鳉jun基因,其蛋白结构含有经典的Jun超家族结构域和bZIP结构域,其蛋白序列进化关系和泛表达模式也符合jun基因的特性。
在突变体青鳉中,在59aa处Jun蛋白翻译终止,导致其完全缺失bZGIP蛋白结构域,这可能是突变体具有抗病毒效果的重要原因。




















 鲁公网安备37020202370212号
鲁公网安备37020202370212号